Male genitalia
See Male Dissections for labelled dissections of the features mentioned here
|
|
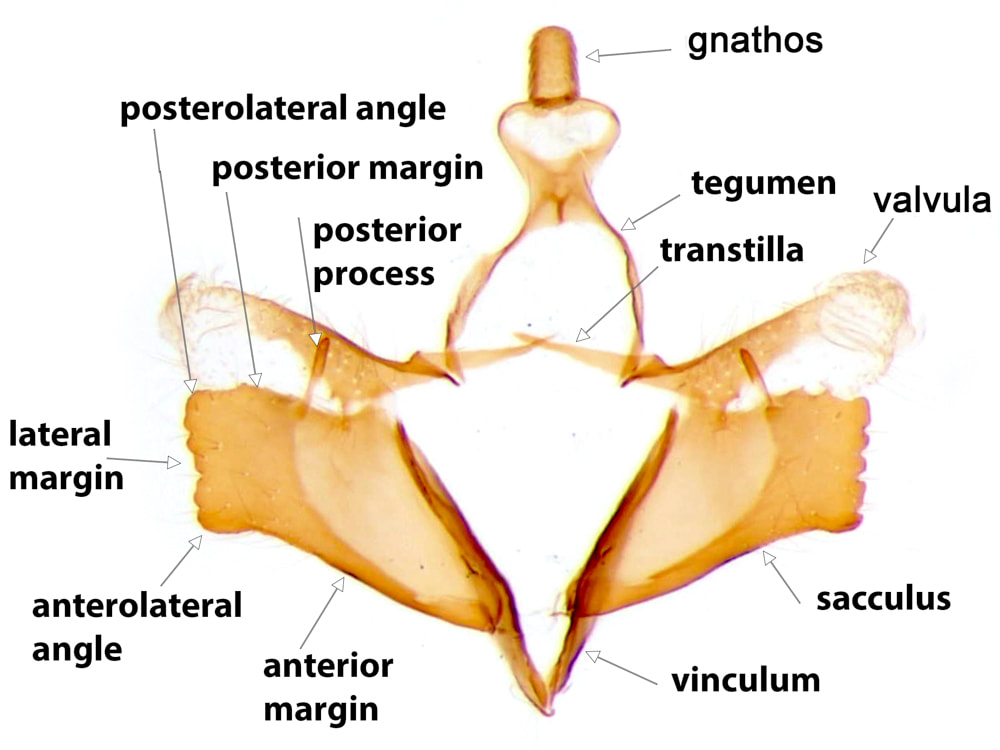
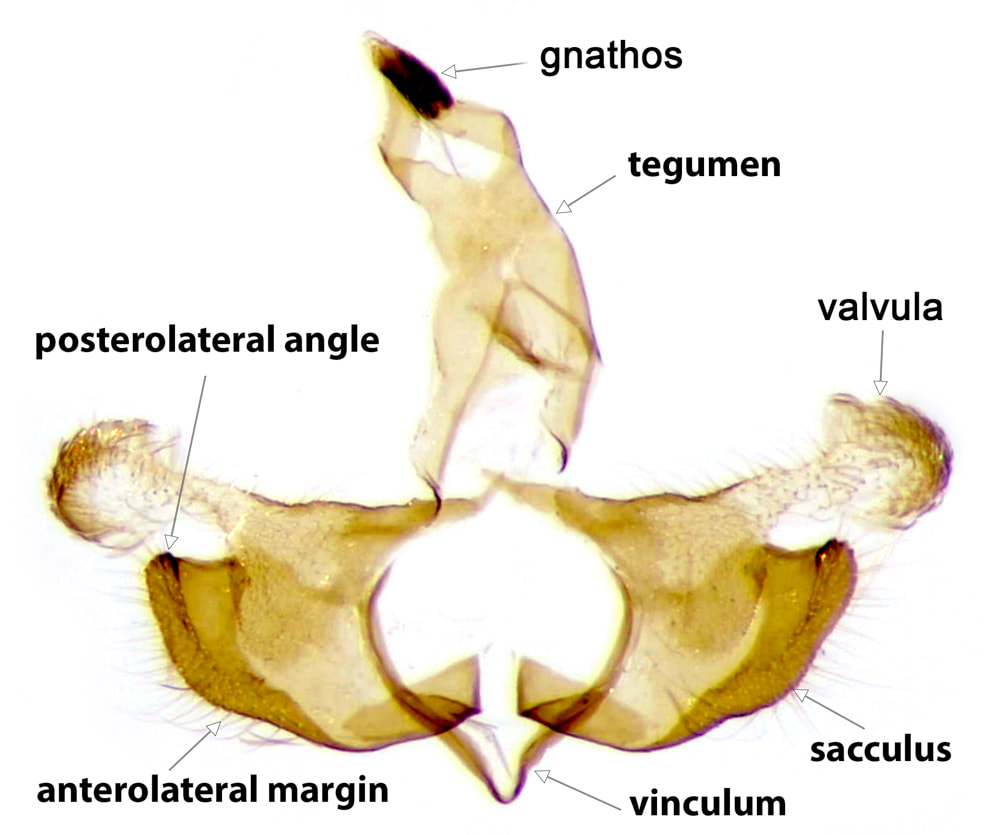
The most important male genital structures used in identification of the species are the sacculus and aedeagus.
The usual layout of Lepidopteran genitalia is based on a ring formed by a posterodorsal tegumen and an anteroventral vinculum. In the Coleophoridae, the vinculum is a narrow well-sclerotised "V" to which the base of the outer surface of the sacculus is attached. The vinculum may be produced somewhat anteriorly and usually dorsally and its ventral margins tethered by a more or less sclerotised flap* - this produced and tethered part of the vinculum is the saccus. There is often a deficiency in the sclerotisation at the apex of the saccus such that the vinculum is effectively composed of a pair of sclerotised bars. The natural shape of the vinculum is seldom preserved in the set position of the usual slide preparation and is not used in identification. In most Lepidopteran families the posterodorsal ends of the vinculum are connected to the anterior ends of the tegumen, forming an articulation. In the Coleophoridae the posterior ends of the vinculum usually have no sclerotised attachments (1)
The tegumen has dorsal and ventral surfaces. The ventral surface is hyaline and so not seen in images that only show the sclerotised parts. The dorsal surface is sclerotised for a variable extent of its posterior portion; anteriorly the sclerotisation is confined to the lateral walls as an arc or "arm" connecting the tegumen to the base of the valval costa on each side. There are two basic forms of tegumen, which I have termed "arched" and "tubular". In an arched tegumen the "arms" are long and narrow and the midline plate is short, extending posteriorly from the point of junction of the two sides of the arch. In a tubular tegumen the midline plate is extensive, clearly longer than the arms. A discrete gnathos** is articulated at the apex of the tegumen. The gnathos is composed of a pair of lateral arms emerging from each side of the posterior margin of the tegumen and a (usually ovoid) median lobe. The lateral arms are directed ventrally and slightly posteriorly, forming an angle of ~110° to the tegumen when viewed laterally in the natural position, and they converge towards the midline. The median lobe is articulated at the apex of these lateral arms and is often retroflexed back so as to overlay the lateral arms and the space between them. The narrowest part of the central lobe is usually at this articulation. The anal tube enters the genital capsule between the dorsal and ventral surfaces of the tegumen and emerges in the space bounded by the posterior margin of the tegumen and the lateral arms of the gnathos.
The Coleophoran valvae are clearly differentiated into a strongly sclerotised anteroventral sacculus and a posterodorsal costa which terminates in a flimsy weakly sclerotised appendage, which I will call the valvula***. The sacculus (pl. sacculi) varies considerably between the species. Its outer surface is attached at its base to the vinculum. In the keys the features of the sacculus are described in their set position with the valvae opened out (note, for example, that the anterior margin of the sacculus in the set position is ventral in the closed genital capsule). Where there are identifiable anterolateral and posterolateral angles, anterior, lateral and usually posterior margins are also identifiable. The anterior margin is usually smoothly curved or straight. The anterolateral angle may be produced into an anterior projection, which may take the form of a more-or-less discrete strongly sclerotised anterior process. The lateral margin may be straight or curved, crenulate, dentate or excavate. The posterolateral angle is often produced into a discrete and sometimes quite elaborate posterior process. Sometimes a posterior process arises from the posterior margin. Where there is no identifiable anterolateral angle there is a continuous anterolateral margin. This may terminate posteriorly at an identifiable posterolateral angle, or laterally as a blunt or pointed lateral projection or lateral process. In a few species the sacculus is reduced to a small triangle or a narrow bar. Where features of the sacculus are notably different in the set than the natural position this is mentioned on the page for the individual species.
The base of the costa is well-sclerotised and usually extends over the internal surface of the valva as a sclerotised plate or raised fold either of which has an extensive connection with the sacculus (this connection is such that the exact boundaries of each structure are poorly defined). The valvula is a flimsy, weakly sclerotised and usually somewhat hairy structure that projects at the lateral end of the costa. The dimensions of the valvula are used in identification of a few species. The medial end of the costa is produced medially into the transtilla (which acts as the posterodorsal support for the aedeagus). In a few species the medial end of the transtilla is elaborated into a distinctive process. The extent of the transtilla varies from a short spur to a complete sclerotised bar fused with its partner in the midline.
The anteroventral support for the aedeagus is provided by a strong band of sclerotisation termed the juxta, which connects the inner surface of the medial ends of the sacculi to the ventral surface of the base of the tunica.
The valvae are connected to the integument by the A8/9 instersegmental membrane. This membrane connected to the ventral surface of the posterior margin of the saccus (such that the saccus itself is anterior and internal to the connection); the outer surface of the sidearms of the vinculum; the base of the costa; and the arch formed by the sidearms and anterior margin of the dorsal plate of the tegumen.
*When this flap is strongly sclerotised and extensive eg C.lutipennella it makes it impossible to open the valvae out for a standard slide prep without disrupting it.
**What has been termed here as the gnathos has routinely been referred to in most reference works as the uncus. For two main reasons I do not accept that this structure is an uncus: Firstly the anal tube opens between the lateral arms and dorsal to the median lobe - the anal tube never opens dorsal to the lepidopteran uncus, but always opens between the gnathos and the ventral surface of the apex of the tegumen (when a gnathos is present). Secondly, it has side arms and a central lobe - an arrangement that is common for a gnathos, but never for an uncus.
***Unfortunately, it is traditional to refer to what I have termed the valvula as the "valva". I think this is confusing, as it leaves us with no unifying term to use for the sacculo-costal complex as a whole. Although use of the term "valvula" has been largely abandoned, I think that in the case of the Coleophoridae it is appropriate and improves clarity of nomenclature.
The usual layout of Lepidopteran genitalia is based on a ring formed by a posterodorsal tegumen and an anteroventral vinculum. In the Coleophoridae, the vinculum is a narrow well-sclerotised "V" to which the base of the outer surface of the sacculus is attached. The vinculum may be produced somewhat anteriorly and usually dorsally and its ventral margins tethered by a more or less sclerotised flap* - this produced and tethered part of the vinculum is the saccus. There is often a deficiency in the sclerotisation at the apex of the saccus such that the vinculum is effectively composed of a pair of sclerotised bars. The natural shape of the vinculum is seldom preserved in the set position of the usual slide preparation and is not used in identification. In most Lepidopteran families the posterodorsal ends of the vinculum are connected to the anterior ends of the tegumen, forming an articulation. In the Coleophoridae the posterior ends of the vinculum usually have no sclerotised attachments (1)
The tegumen has dorsal and ventral surfaces. The ventral surface is hyaline and so not seen in images that only show the sclerotised parts. The dorsal surface is sclerotised for a variable extent of its posterior portion; anteriorly the sclerotisation is confined to the lateral walls as an arc or "arm" connecting the tegumen to the base of the valval costa on each side. There are two basic forms of tegumen, which I have termed "arched" and "tubular". In an arched tegumen the "arms" are long and narrow and the midline plate is short, extending posteriorly from the point of junction of the two sides of the arch. In a tubular tegumen the midline plate is extensive, clearly longer than the arms. A discrete gnathos** is articulated at the apex of the tegumen. The gnathos is composed of a pair of lateral arms emerging from each side of the posterior margin of the tegumen and a (usually ovoid) median lobe. The lateral arms are directed ventrally and slightly posteriorly, forming an angle of ~110° to the tegumen when viewed laterally in the natural position, and they converge towards the midline. The median lobe is articulated at the apex of these lateral arms and is often retroflexed back so as to overlay the lateral arms and the space between them. The narrowest part of the central lobe is usually at this articulation. The anal tube enters the genital capsule between the dorsal and ventral surfaces of the tegumen and emerges in the space bounded by the posterior margin of the tegumen and the lateral arms of the gnathos.
The Coleophoran valvae are clearly differentiated into a strongly sclerotised anteroventral sacculus and a posterodorsal costa which terminates in a flimsy weakly sclerotised appendage, which I will call the valvula***. The sacculus (pl. sacculi) varies considerably between the species. Its outer surface is attached at its base to the vinculum. In the keys the features of the sacculus are described in their set position with the valvae opened out (note, for example, that the anterior margin of the sacculus in the set position is ventral in the closed genital capsule). Where there are identifiable anterolateral and posterolateral angles, anterior, lateral and usually posterior margins are also identifiable. The anterior margin is usually smoothly curved or straight. The anterolateral angle may be produced into an anterior projection, which may take the form of a more-or-less discrete strongly sclerotised anterior process. The lateral margin may be straight or curved, crenulate, dentate or excavate. The posterolateral angle is often produced into a discrete and sometimes quite elaborate posterior process. Sometimes a posterior process arises from the posterior margin. Where there is no identifiable anterolateral angle there is a continuous anterolateral margin. This may terminate posteriorly at an identifiable posterolateral angle, or laterally as a blunt or pointed lateral projection or lateral process. In a few species the sacculus is reduced to a small triangle or a narrow bar. Where features of the sacculus are notably different in the set than the natural position this is mentioned on the page for the individual species.
The base of the costa is well-sclerotised and usually extends over the internal surface of the valva as a sclerotised plate or raised fold either of which has an extensive connection with the sacculus (this connection is such that the exact boundaries of each structure are poorly defined). The valvula is a flimsy, weakly sclerotised and usually somewhat hairy structure that projects at the lateral end of the costa. The dimensions of the valvula are used in identification of a few species. The medial end of the costa is produced medially into the transtilla (which acts as the posterodorsal support for the aedeagus). In a few species the medial end of the transtilla is elaborated into a distinctive process. The extent of the transtilla varies from a short spur to a complete sclerotised bar fused with its partner in the midline.
The anteroventral support for the aedeagus is provided by a strong band of sclerotisation termed the juxta, which connects the inner surface of the medial ends of the sacculi to the ventral surface of the base of the tunica.
The valvae are connected to the integument by the A8/9 instersegmental membrane. This membrane connected to the ventral surface of the posterior margin of the saccus (such that the saccus itself is anterior and internal to the connection); the outer surface of the sidearms of the vinculum; the base of the costa; and the arch formed by the sidearms and anterior margin of the dorsal plate of the tegumen.
*When this flap is strongly sclerotised and extensive eg C.lutipennella it makes it impossible to open the valvae out for a standard slide prep without disrupting it.
**What has been termed here as the gnathos has routinely been referred to in most reference works as the uncus. For two main reasons I do not accept that this structure is an uncus: Firstly the anal tube opens between the lateral arms and dorsal to the median lobe - the anal tube never opens dorsal to the lepidopteran uncus, but always opens between the gnathos and the ventral surface of the apex of the tegumen (when a gnathos is present). Secondly, it has side arms and a central lobe - an arrangement that is common for a gnathos, but never for an uncus.
***Unfortunately, it is traditional to refer to what I have termed the valvula as the "valva". I think this is confusing, as it leaves us with no unifying term to use for the sacculo-costal complex as a whole. Although use of the term "valvula" has been largely abandoned, I think that in the case of the Coleophoridae it is appropriate and improves clarity of nomenclature.
|
|
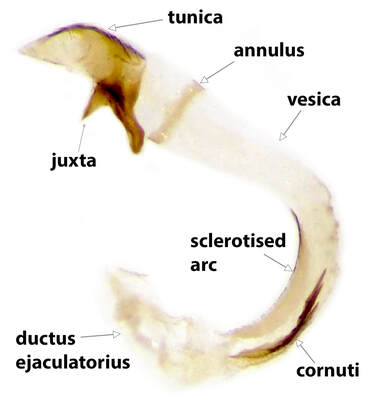
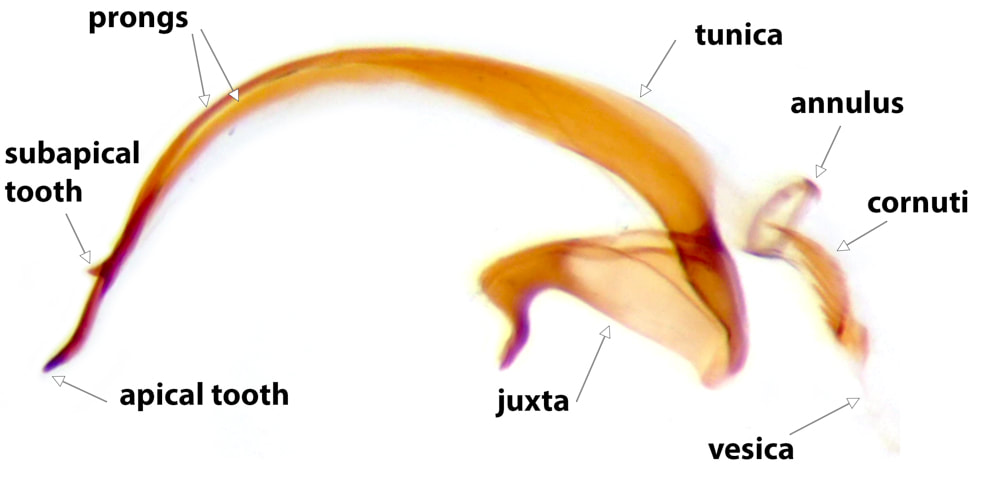
There are broadly two main varieties of Coleophoran aedeagus: one in which the tunica is a simple sheath, the other in which it is divided into 2 prongs. Prongs may have teeth, usually on the dorsal surface and which may be apical, subapical or midway along one or both prongs. (As the juxta is fused to the tunica it should be carefully detached from the sacculi when the aedeagus is removed). In most Lepidopteran genitalia the vesica is entirely contained within the tunica, but in Coleophoran genitalia it extends proximally from the tunica as a curved, rigid, (mostly) hyaline, tube. Just proximal to the tunica this tube has a sclerotised ring in its wall, the annulus. It is convenient, and conventional, (though probably not anatomically accurate), to term the sclerotised apex of the aedeagus as the tunica and the hyaline tube as the vesica. Cornuti, when present, are nearly always contained within the vesica. The concave side of the curve of the vesica is usually sclerotised to some extent. The vesica is continuous proximally with the ductus ejaculatorius; the junction between the two occurring where the walls of the tube lose rigidity.
(1) In C.limoniella the posterior ends of the vinculum do have a sclerotised articulation with the anterior ends of the tegumen
